www.MUTAVI.info
31th of July 2011
Red-and black-headed Gouldian Finch; evolution at work?
The Netherlands
Email: dhschamhart@hotmail.com
After its discovery (about 200 years ago) there has been much debate about whether the Gouldian Finch with its three different head colors should be considered as one or three , separate species. Today, it is generally accepted as a single species. Yet, it turns out, that those ornithologists assuming three species were not entirely wrong.,
The Gouldian Finch (Erythrura gouldiae) has raised considerable interest within the community of evolutionary biologists, certainly and understandably those in Australia. With this species S. Pryke and her colleagues, at Macquarie University - Sydney, are studying the mechanism of evolution of new species. It was long thought that speciation depends on a physical separation between the daughter and the parental population by a geological barrier such as a mountain range, desert, ocean, or climatic condition (allopatric speciation). Eventually, after thousands to millions of years, the daughter and parental populations differ to such an extent that they no longer can interbreed - two new species have emerged. More recently, computer models and experiments with various animals and plants suggest that a new species may also emerge within the same geographic area in which a geological barrier does not play a role (sympatric speciation).
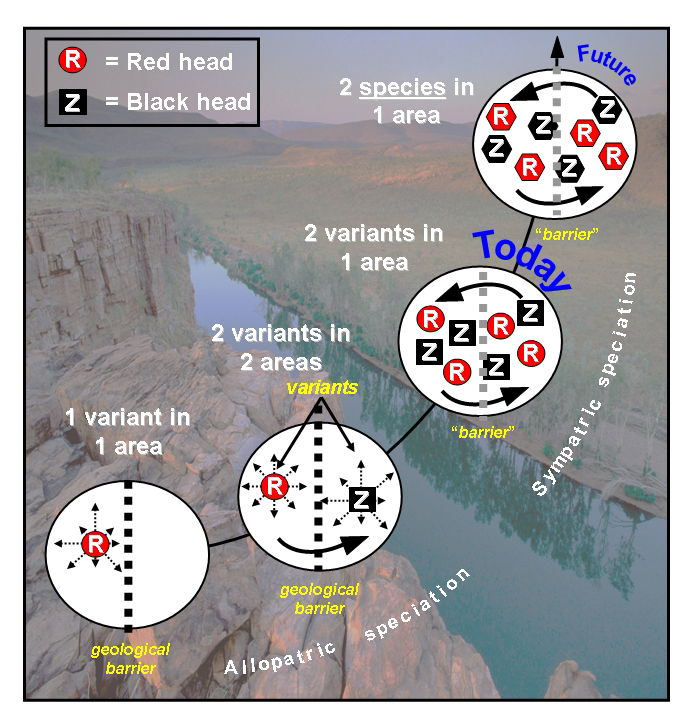
Fig. 1. Possible evolution of red and black head Gouldian Finch variants. Whether the variants in the (distant) future will evolve into two species remains to be seen.
The latter mode of speciation, without a geological barrier, now seems to occur with the Gouldian Finch (Fig.1). In the wild, the three color morphs coexist together throughout their entire geographic range at relative stable frequencies: black head (70-80%), red heads (20-30%) and yellow heads (less than 0.1%). It is believed that thousands to millions of years ago, the black head originated as a mutant of the parental red head (1). Initially, the black head remained in existence, not as a new species, but as a genetically different variant, possibly separated from the red head by two plateaus (Carpentarian and Kimberley Plateau-Arnhem Land Barrier, Australia) (2 variants in two areas, Fig. 1) (2,3). Later, the two variants came back into contact and to this day coexist together in the same geographical area (2 variants in one area, Fig. 1). This could mean that currently the Gouldian Finch variants represent an intermediate stage towards the formation of two new species. However, the crucial question is: How it is that red and black heads coexist and do not merge into a homogeneous population of (almost) entirely red heads? Indeed, interbreeding of red and black heads produces fertile offspring and mixed morph breeding in the wild is not uncommon (up to 30%) (4). Moreover, the locus for red is sex, Z- chromosome linked and dominant. Consequently, the offspring of mixed, homozygous pairs will, taken together, consist of many (75%) phenotypical red heads and few black heads. So, ultimate evolution into two, mutually infertile species requires mechanisms to keep these coexisting variants segregated. For this, both prezygotic and postzygotic mechanisms have been described, encompassing sexual isolation (e.g. non-random mating, behavioral factors) and sterile/non-viable offspring.
The Australian researchers are exploring the (still limited) isolation between the Gouldian Finch variants studying both behavioral and genetic factors.
Behavioral Factors. (1) It was found that red heads (RH) are very aggressive against black heads (BH) (and orange heads) at e.g. foraging and occupying the best nesting sites (Fig. 2) (4-8). (2) Females showed a clear mating preference for a partner with the same head color (assortative mating). In the Gouldian Finch this preference appears to be genetically determined and not, as often assumed, a result of imprinting (acquired) (8).
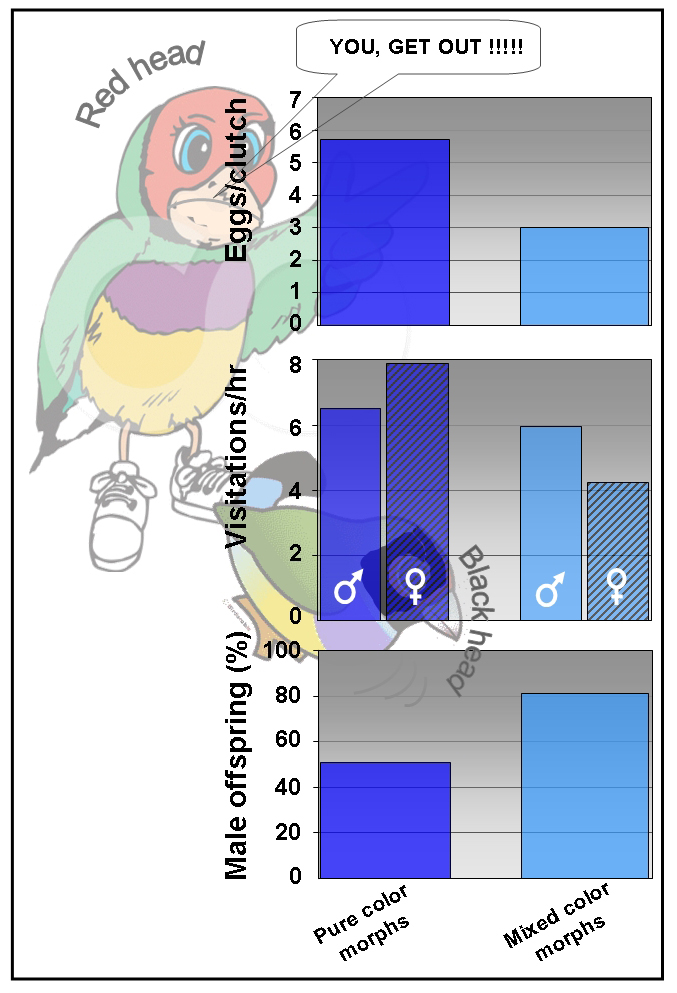
Fig. 2. Behavior, number of eggs per clutch, visitations (nest visits per hour) by male and female, and percentage of males per clutch with phenotypically compatible (RH x RH, BH x BH pure color morphs) and incompatible (RH x BH, BH x RH mixed color morphs) pairs.
(3) The life history theory is a theory from evolutionary biology that addresses among others the extent of investment/energy by a female in reproduction vs. her opinion about certain environmental conditions, such as food, and the quality of the male partner (dominance, attractiveness, health) (5,8-10). If breeding with a high-quality male is not possible and the female breeds with a low-quality male, she will spend less effort in raising the offspring. Moreover, for birds it was suggested that female birds can bias the gender of their clutch or in other words influence the division sons/daughters (sex ratio). Until recently there was little experimental evidence in birds for this theory. However, in 2009 Pryke and Griffith working with over 200 pairs of Gouldian Finches consisting of phenotypically (appearance) compatible and incompatible pairs of red and black heads convincingly demonstrated the determinative role of the female in the upbringing of the offspring (11). Compared with phenotypically compatible pairs (RH x RH or BH x BH), crosses of incompatible pairs, RH x BH or BH x RH, resulted in less eggs per clutch (average 3 instead of 6) and a limited feeding effort (nest visits per hour) of the female (Fig. 2). Feeding by the male, however, was similar in both compatible and incompatible pairs.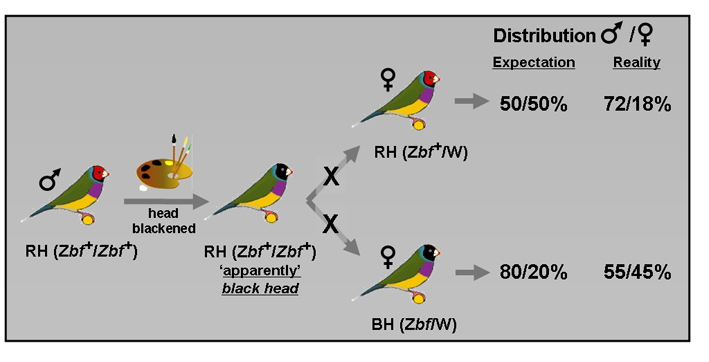
Fig. 3. Crosses between black-dyed RH male x RH female (genetically compatible, but phenotypically, apparently incompatible) (top) and black-dyed RH male x BH female (genetically incompatible, but phenotypically appropriate) (bottom). These experiments showed that despite genetic (in) compatibility the female can affect the ratio of male/female eggs, led by the head color. The genetic notation is that described by Van den Abeele (12).
It also appeared that Gouldian Finch females have the ability to adjust the sex of their eggs (Fig. 2). This fascinating ability of the female Gouldian Finches was derived from experimental crosses between non-manipulated females (RH and BH) and red head males of which the head was blackened (11). The offspring distribution male/female of two types of crosses was determined: (1) black-dyed red head (RH-black-dyed) male x RH female (genetically compatible, but phenotypically incompatible) and (2) black-dyed red head male x BH female (genetically incompatible, but phenotypically compatible) (Fig. 3). In genetically compatible, but phenotypically incompatible pairings (RH-black-dyed male x RH female) the clutches consisted of 72% males, contrary to the genetically expected 50%. By contrast, the clutches of black-dyed RH male x BH female, an apparently equal match, consisted of 55% males. With the latter, genetic incompatible cross, the genetic expectation was 80% males (see below). It's like black-dyed RH x BH is genetically compatible. These results demonstrate that the female can affect the distribution male/female eggs (sex ratio) within a clutch. As a possible explanation for the relative low female offspring of phenotypically incompatible pairings it is suggested that in the wild the survival of female offspring is lower than males. The cost/benefit for the female of an incompatible pairing would be too high, in particular for raising daughters (life history theory).Taken together, the ability to assortative mating, to adjust the sex of their eggs and to allocate resources hamper mating between different color morphs and ultimately may prevent complete mixing of red and black heads. The Gouldian Finch female has much more influence over the outcome of reproduction than has previously been recognized.
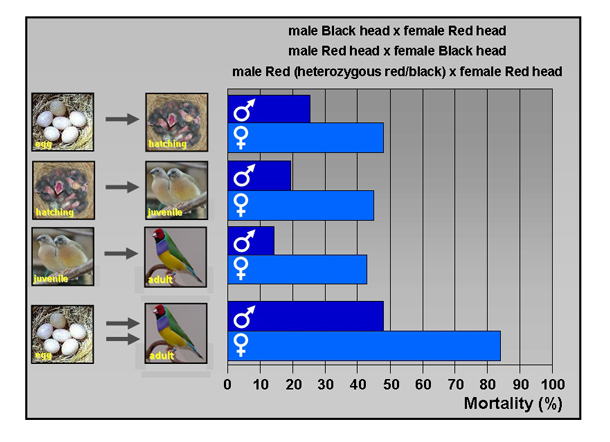
Fig. 4. Mortality among offspring of genetically incompatible (BH x RH and RH x BH) and phenotypically compatible, but genotypically heterozygous red/black head male, Zbf+/Zbf, x RH female in each of the first phases of life, and the total (from egg to adult). Gender (in egg and before adult) was determined using a polymerase chain reaction (PCR).
Genetic factors. Pyrke and coworkers extended their investigation of the isolation between red and black heads, including genetic factors (2). Of crosses between genetically incompatible pairs (RH x BH and BH x RH), the mortality of male and female offspring in each of the first 3 phases of life was studied. These phases were: (1) from (fertilized) egg to hatching, (2) from chicken to juvenile in foster nest (60 days), and (3) juvenile to maturity (140 days) (Fig. 4). During each of the individual phases approximately 20% of male and 46% of female offspring died. Compared with compatible crosses (RH x RH and BH x BH), this means a 40% and 83% higher mortality of male and female offspring respectively during complete development (from egg to adult). Interestingly, crosses between phenotypically compatible, but genetically heterozygous red/black males and red head females resulted in a similar low survival of offspring (Fig. 4). These observations and in particular the large difference in viability between the male and female offspring of incompatible pairs indicate a genetic incompatibility or genetically determined adverse effect on viability.
The latter has a strong resemblance to crosses between different species. Already in 1922, Haldane suggested that when in the offspring of two different animal and plant species one sex is absent, rare, or sterile, that sex is the heterozygous sex (X/Y or Z/W) (Handan's rule) (13,14). For birds, an example of Haldane's rule is the among canary breeders well-known cross between the Red Hooded Siskin and the Canary. To my knowledge, no fertile female offspring will arise from this cross, but only - sparse - fertile male hybrids. With respect to viability the explanation of Haldane's rule seems to be related to Z-linked, recessive alleles with adverse effects in hybrids. The effects of recessive deleterious alleles in the homogametic hybrids (bird males, Z/Z) will be masked at least in part by alleles from the other parent. In the heterogametic hybrids (bird females, Z/W), there is no masking and these deleterious alleles will manifest their full effects. Note that these alleles are deleterious and recessive only in the hybrid genetic background. In the pure species background, they are either neutral or advantageous, and their dominance relations are irrelevant.
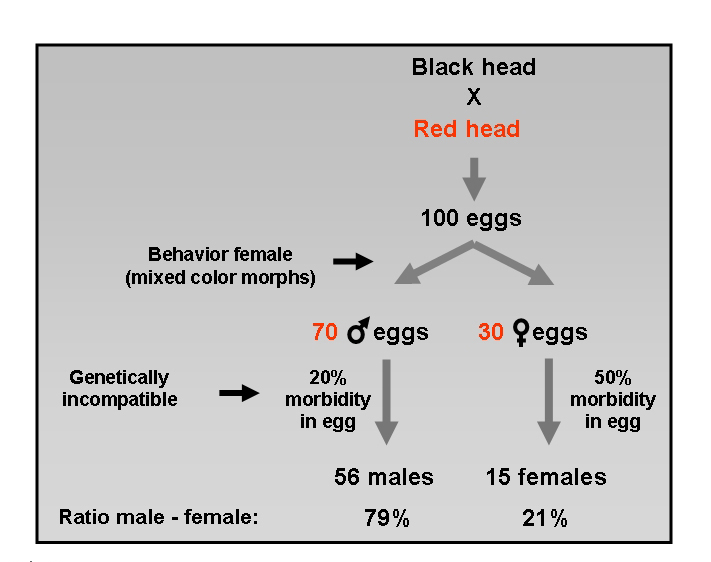
Fig. 5. An example explaining the realization of the distribution male/female (and survival) at hatching from an incompatible pair. Note: after hatching morbidity among especially the female offspring will still grow strongly (Fig. 4).
How about the Gouldian Finch? The high percentage (80%) mortality among the female offspring of genetically incompatible pairs seems in accord with Haldane's rule for the viability of hybrids. Figure 5 is a calculation example of how the distribution (and viability) of male-female of a cross BH x RH accumulates due to the various phenotypical and genotypical factors. These data suggest major genetic divergence between red- and black-headed Gouldian Finches.
All these results, the behavioral factors and genetic factors now seem to justify the conclusion that the red- and black-headed Gouldian Finch should be considered as intermediate species and the offspring as intermediate hybrids.
These observations and conclusions are not only of importance within the context of basic evolutionary biology, but also within the proposed program for the release of Gouldian Finches, as an endangered species, in the wild. It should be ensured that only pure-bred birds will be used. So, certainly the release of heterozygous red/black head males should be avoided (Fig. 4).
Considering the breeder of Gouldian Finches, he/she should be aware that breeding with Gouldian Finches is breeding with two variants (subspecies?). Not only the viability of the offspring from the cross red x black head will on the average be low, but such a (forced) cross seems to be contrary to a natural process currently happening in the Australian wild. The (fundamental) question arises whether the cross red x black head can still be considered as natural breeding.
Finally, it should be noted that it is unknown to what extent female preference (and consequently the behavior: number of eggs, sex ratio, etc.) are affected by other, not naturally occurring type of color mutants (for example, blue or white breasted). However, genetic considerations (Haldanes rule) predicts that the viability among the offspring (especially of females) of a cross such as male black head/ white breast/ green back x female red head/purple breast/green back will be low and, including possible behavioral aspects, as low as the wildtype male black head x wildtype red head female. It would of interest to explore this aspect in cooperative effort.
Literature:
A summary of many of the key articles can be found at:
http://www.savethegouldian.org/Research.html
[1] Van den Abeele, D. 2002.
De gouldamadine eens anders bekeken.
MUTAVI, Research & Advice group.
[2] Pryke, S. R. & Griffith, S. C. 2009.
Postzygotic genetic incompatibility between sympatric color morphs.
Evolution 63: 793-798.
[3] Jennings, W. B. & Edwards, S. V. 2005.
Speciational history of Australian grass finches (poephila) inferred from thirty
gene trees,
Evolution 59: 2033-2047.
[4] Pryke, S. R. 2007.
Fiery red heads: female dominance among head color morphs in the Gouldian finch.
Behavioral Ecology 18: 621-627.
[5] Pryke, S. R. & Griffith, S. C. 2009.
Socially mediated trade-offs between aggression and parental effort in competing
color morphs.
American Naturalist 174: 455-464.
[6] Pryke, S. R. 2009.
Is red an innate or learned signal of aggression and intimidation?
Animal Behaviour 78: 393-398.
[7] Pryke, S. R. & Griffith, S. C. 2006.
Red dominates black: Agonistic signalling among head morphs in the colour polymorphic
Gouldian finch.
Proceedings of the Royal Society of London B. 273: 949-957.
[8] Pryke, S. R. 2010.
Sex chromosome linkage of mate preferences and color signal maintains assortative mating
between interbreeding finch morphs.
Evolution 65: 1301-1310.
[9] Pryke, S. R. & Griffith, S. C. 2010.
Maternal adjustment of parental effort in relation to mate compatibility affects
offspring development.
Behavioral Ecology 21: 226-232.
[10] Martin, T.E. 2004.
Avian life-history evolution has an eminent past: Does it have a bright future?
The Auk 121: 289-301.
[11] Pryke, S. R. & Griffith, S. C. 2009.
Genetic incompatibility drives sex allocation and maternal investment
in a polymorphic finch.
Science 323: 1605-1607.
[12] Van den Abeele, D. 2010.
De gouldamadine eens anders bekeken. Deel 2.
MUTAVI, Research & Advice group
[13] Graves, J.A.M. & O'Neill, W. 1997.
Sex Chromosome Evolution and Haldane's Rule J.
Heredity 359: 738:358-360.
[14] Turelli, M. & Orr, H.A. 1995.
The dominance theory of Haldanes rule.
Genetics 140: 389-402.
© 2003 - Inte Onsman - MUTAVI Research & Advice Group
WebDesign © 2007 - Martin Rasek



