www.MUTAVI.info
29th of December 2003
The Involvement of Recessive Pied in the Origin of the Dark Eyed Clears in the Budgerigar
[Melopsittacus undulatus]
MUTAVI
Research & Advice Group
Recessive pied
The first pied mutation in Budgerigars was established in 1932. The mutation appeared to be
recessive and was called Danish pied. These birds were also referred to as Finnish pieds for a
short time because the first species who reached England came from Finland
[17].
The very first mutant was discovered in a mixed coloured flock of Budgerigars breeding as a
colony. Therefore is was not possible to find out from which parents this bird was derived.
One of the first aviculturists who understood that this actually was an important new mutation
was C. af Enehjelm, head of the Helsinki zoo in Finland. Thanks to his interest and dedication
this mutation was quickly distributed through the whole of Europe.
During World war II Danish pieds almost died out but the Danish fancier Walter Langberg
saved the mutation from total extinction.
Dutch pied
The source of the first Dutch pieds cannot be established for sure. In "The Handbook for
Budgerigar Breeders" written by W.Beckmann (1966) is written that Mr. Van Dijk, head of
"Animali" zoo in Eindhoven (The Netherlands) was already breeding Dutch pieds before
World war II. Also foreign sources mention the year 1939 to be the year in which the first
Dutch pieds appeared and even in America the mutation is known as "Dutch pied".
Many authors consider the clearflight to be a selective form of Dutch pied and both in
England and America these clearflights are known as "continental clearflights".
The first specimens were developed by R. Raemaker from Bruxelles who claimed it took him
several years to breed the first clearflights from birds who initially had only a few
unpigmented feathers.
During the late Forties and the early Fifties, people from Europe mentioned black-eyed
"albinos" and "lutinos". During several years it was presumed that these birds could only be
bred by cross breeding them, but later on evidence was found that this was not true.
Experimental matings with these birds adventually cleared the matter. They were in
fact a combination of recessive pied and continental clearflights (a selective form of Dutch pied).
These birds also did not have black eyes but the plum eyes of the recessive pieds.
Yet they were known as dark eyed clears.
Unfortunately Dutch pieds never became very popular in the Netherlands
unlike America and Germany where some fanciers breed them with great enthousiasm
[22,23,24].
There are some remarkable similarities between Dutch pieds and recessive pieds to be found
in literature. If a DF Dutch pied is mated to a normal, one expects 100% Dutch pieds.
However, some authors report the appearance of a single normal youngster among the
offspring of such mating (...).
Male Dutch pieds show more pied feathers than female Dutch pieds. This is also the case in
Australian pieds and recessive Danisch pieds. In the USA and in England one distinguishes
Dutch pieds from continental clearflights. Some people state that continental clearflights
could be a selective form of Dutch pied and having seen my own results with Dutch pieds I
am willing to believe that this is actually the case. The main question is why the combination
Danish- / Dutch pied produces a dark eyed clear bird and the combination Danish
/ Australian does not. To find the answer to this question, we have to return to the early
embryonical state and the source where melanoblasts, the precursors of melanocytes, are
produced.
The research
The origin of pigmentation in vertebrates has always been a favourite subject of scientists all
over the world
[7,9,10,11,15]. The first authors were very much impressed by the starshaped cells
containing melanin they found in the skin.
Already in 1860 Kölliker described migration of pigment cells in the skin which were derived
from mesenchymal (brain) tissue. This point of view was already confirmed by Kerbert in
1877 for reptiles and birds, again for humans and birds by Riehl in 1885 and several
mammals and again birds by Ehrmann. In 1890 Haecker studied pigmentation in bird feathers
and came to the conclusion that feather pigmentation is caused by migrating pigment cells
derived from mesenchymal sources. (The mesenchym is a network made of embryonal tissue
from which in a later state the ordinary tissues, blood and lymfe vessels are derived.)
Soon it became clear that the source of pigment cells could not be found under the
microscope because melanoblasts (pigment cell precursors) are not to be distinguished from
other cells. Experimental transplants involving amphibians showed notable different
conclusions, for pigment cells do not derive from mesenchymal tissue, or as some people
stated, in the skin itself, but derive from the neural crest. In 1935 this was definitely
confirmed by Dushane
[3] who did research on transplanted parts of the neural crest from
Ambystoma (a Mexican salamander).
In the fowl this problem was first investigated in 1936 and 1939 by Dorris
[2] and also in 1941 by Ris
[18].
By studying cultivated tissues and transplantation experiments, they concluded that the neural crest was able to
produce pigment cells and that the mesoderm as well as the skin were not able to do so.
The neural crest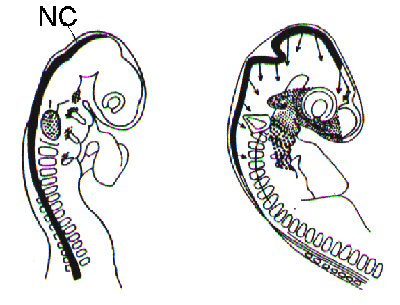
The neural crest in fowl embryos was first described by His in 1868.
In a very early stage the
neural walls are formed. These neural walls rise and close in order to form the neural tube,
the precursor of the central nerve system and the spinal cord. The neural crest is situated on
the backside of the neural tube and immediately starts dispersing celprecursors.
Along both sides of the neural tube somites are to be found. Somites are blockshaped
embryonic tissue elements which turn into a dermatome from which skin segments are
formed. All skin segments together eventually will form the entire skin.
Pied genes
Genes causing nonuniform pigmentation patterns have been found and investigated in many vertebrates. In
mice more than fifteen different pied mutations are known, some of them are even multiple
allelic.
Pied genes are able to manifest themselves in different ways as many research projects that
have been done over the past years have shown. Deol investigated the role of the tissue
environment in the expression of pied genes in mice
[1] and found that some pied genes
affect melanoblasts (pigment cell precursors). Others affect the tissue environment in which
the melanoblasts reside.
During this research particularly interest was taken in the eye
pigmentation of several piebald mutations. Some of these mutations affect eye pigmentation
and others do not.
In the fowl the migratory behaviour of neural crest cells was studied extensively by Noden
[14]. He found that the migratory behaviour of cells derived from the neural crest, is
influenced dramatically by certain interactions with their environment. All neural crest cells,
with regard to which part they came from, do possess the ability to recognize specific
genetical signals, react on them and so determine the direction of their migration.
In 1982 Hirobe published a paper in which he described investigations on the regulation of
melanoblast / melanocyte populations in the skin of newborn mice
[5].
He came to the conclusion that the distribution of melanoblasts / melanocytes into the skin, is
under control of a number of (up to now) unknown semi dominant genes. In 1992 he
publishes the results of a follow-up where he investigated the proliferation and differentiation
of melanocytes
[6] and announced research on the number of genes involved.
Eye pigmentation
The research of Deol inspired me some years ago to compare eye pigmentation from Danish- and Australian
pieds using a light microscope.
The following items are important to know in this respect.
Melanocytes or pigment cells derive from three different sources:
1) The neural crest from where they migrate to all parts of the body including the choroid and
the outher layer of the iris of the eye.
2) The outher ridge of the optic cup which eventually will form the pigment epithelium of the
retina.
3) The neural tube from where neuromelanins are formed who mainly are to be found in
brain tissues.
Summarized the melanocytes of the eye have two sources. To be precise, the melanocytes of
the choroid and the iris stroma (frontside of the iris) are derived from the neural crest, the
melanocytes of the pigment epithelium (retina) and the inner (back) side of the iris are
derived from the edge of the optic cup. The optic cup is the precursed state of the eye, the
definite eyeball and lens are yet to be formed. The pigment epithelium excists of honeycomb
shaped highly specialized melanocytes, residing between the retina and the choroid. The
melanocytes of the choroid reside amongst the bloodvessels of this layer which is situated
between the pigment epithelium and the scleral cartilage (outher layer of the eye in birds).
The difference between eye melanocytes and epidermal (skin) melanocytes is that the
melanocytes of the eye do not disperse their pigment granules into neighbouring tissues
unlike skin melanocytes. The results of my investigations showed that the eyes of Danish
pieds do have unpigmented areas in the choroid, and the outher layer (frontside) of the iris
almost totally lacks pigment cells. The retinal pigment epithelium and the inner layer
(backside) of the iris are unaffected.
The eyes of Australian pieds were completely unaffected and eyes from Dutch pieds have not
yet been investigated but are presumed to be unaffected as well.
Photographs of cross sections
(for best results use a colorprinter and glossy paper)
Recessive pied pigment pattern - Recessive pied wing pattern
Internal pigmentation of the eyes in recessive pied (s / s) birds
Conclusion:
Having seen these results I came to the conclusion that in Danish pieds, exclusively
melanocytes derived from the neural crest are affected by the s-locus. This suggest a possible
defect of the neural crest itself causing a disturbed production of too few, abnormal and also
normal pigment cell precursors. Obviously the s-locus (Danish pied) acts in the neural crest
and not in the skin. In Australian pieds, the Pb-locus could act in the former mentioned
dermatomes or in certain skin areas in which as a result melanoblasts are unable to
differentiate into melanocytes leaving those areas unpigmented.
The Dutch pied gene (Pi-locus) could affect e.g. the proliferative capacities of
melanoblasts because often wingtops, being most far apart from the neural crest, show pied
areas. It could also be possible that the Dutch pied locus is involved in determining the
direction of the migrating melanoblasts.
The real dark eyed clears (or should we say plum-eyed) can only be
achieved by the interaction of Danish- and Dutch pied. Apparently these two loci are playing
a major role in pigment distribution in Budgerigars. The combination Australian- / Dutch
pied will never develop a dark eyed clear phenotype, that is quite obvious.
The mottle is deliberately disregarded in this article, because the cause of the mottled
phenotype differs significant from the other pied mutations.
Breeding Dutch pieds should be recommanded strongly. They are very attractive in
appearance, have bright colours just as Danish pieds and body size is easy to improve.
[1] Deol M.S.,(1973)
The Role of the Tissue Environment in the Expression of Spotting Genes
in the Mouse
Journ.Embryol.Exp.Morphol. Vol.30 no.2; p.p.483-489
[2] Dorris F.,(1939)
The Production of Pigment by Chick Neural Crest in Grafts
to the 3-day Limb Bud
Journal of Exp. Zool. Vol.80; p.p.315-345
[3] Dushane G.P.,(1935)
An Experimental Study of the Origin of Pigment Cells in Amphibia
Journal of Exp.Zool.Vol.72,no.1; p.p.1-31
[4] Dushane G.P.,(1943)
The Embryology of Vertebrate Pigment Cells. Part I. Amphibia
Quarterly Review of Biology Vol.18,no.2; p.p.109-127
[5] Hirobe T.,(1982)
Genes Involved in Regulating the Melanocyte and Melanoblast-
Melanocyte Populations in the Epidermis of Newborn Mouse Skin
Journ. of Exp. Zool. Vol.223; p.p.257-264
[6] Hirobe T.,(1992)
Control of Melanocyte Proliferation and Differentiation in the Mouse
Epidermis
Pigment Cell Research no.5; p.p.1-11
[7] Hulley P.A., Stander C.S., Kidson S.H.,(1991)
Terminal Migration and Early Differentiation of Melanocytes in
Embryonic Chick Skin
Dev.Biol.Vol.145; p.p.182-194
[8] LeDouarin N.M.,(1982)
The Neural Crest (Book); p.p.216-248
[9] LeDouarin N.M.,(1982)
Pigment Cells
The Neural Crest (Cambr.Univ.Press); p.p.108-133
[10]LeDouarin N.M.,(1984)
Cell Migrations in Embryos
Cell Vol.38; p.p.353-360
[11]Lerner A.B.,(1980)
Behavior of Pigment Cells
Journ.Invest.Derm. Vol.75; p.p.121
[12]Marusich M.F., Weston J.A.,(1991)
Development of the Neural Crest
Curr.Op.in Genet. and Dev. Vol.1; p.p.221-229
[13]Mayer T.C.,(1977)
Enhancement of Melanocyte Development from Piebald Neural Crest by a
Favorable Tissue Environment
Dev.Biol. Vol.56; p.p.255-262
[14]Noden D.M.,(1975)
An Analysis of the Migratory Behaviour of Avian Cephalic
Neural Crest Cells
Dev.Biol. Vol.42; p.p.106-130
[15]Okun M.R.,(1965)
Histogenesis of Melanocytes
Journ.Invest.Derm. Vol.44 no.5; p.p.285-299
[16]Rawles M.E.,(1947)
Origin of Pigment Cells from the Neural Crest in the Mouse Embryo
Physiol.Zoology Vol.20; p.p.248-266
[17]Rogers C.,(1987)
The World of Budgerigars: p.p.73-81
Nimrod Press LTD, Engeland
[18]Ris H.,(1941)
An Experimental Study on the Origin of Melanophores in Birds
Physiol.Zoology Vol.14; p.p.48-69
[19]Stern C.D., Artinger K.B., Bronner-Fraser M.,(1991)
Tissue Interactions Affecting the Migration and Differentiation
of Neural Crest Cells in the Chick Embryo
Development Vol.113; p.p.207-216
[20]Taylor T.G., Warner C.,(1986)
Genetics for Budgerigar Breeders: p.p.90-94
The Budgerigar Society, Engeland
[21]Thiery J.P., Duband J.L., Delouvee A.,(1982)
Pathways and Mechanisms of Avian Trunk Neural Crest
Cell Migration and Localization
Dev.Biol. Vol.93; p.p.324-343
[22]Travnicek R.,(1986)
The Death of Beauty
Budgerigar World, no.7: p.p.15-16
[23]Versluis A.,(1993)
Zwartogen
Budgie 4e jaargang no.6: p.p.17-23
[24]Yorke K.,(1993)
Budgerigar World, no.2: p.p.25-26
© 2003 - Inte Onsman - MUTAVI Research & Advice Group
WebDesign © 2007 - Martin Rasek



